





Mezofil tejsavtermelő baktériumok-száma
Pontos definíció nem létezik a tejsavbaktérium (angolul: Lactic Acid Bacteria, LAB) kifejezésre, amely nem rendszertani kategória, hanem közös anyagcsere- és élettani sajátosságokkal rendelkező baktériumcsoportok gyűjtőneve. A tejsavbaktériumokhoz tartozó nemzetségeket az Eubacteria birodalmon belül, a Gram-pozitív baktériumok Firmicutes törzsében találjuk. Az ide tartozó baktériumok – az atipikus sejtfallal rendelkező csoportoktól eltekintve – mind Gram-pozitív módon festődnek és kis guanin+citozin (G+C) tartalommal rendelkeznek (a DNS G+C aránya 50 mol% alatti).
Említést kell tenni a számos hasonló tulajdonságuk miatt gyakorlati szempontból és hagyományosan is a tejsavbaktériumokkal együtt tárgyalt és probiotikus tulajdonságokkal rendelkező bifidobaktériumokról, amelyek filogenetikailag teljesen elkülönülnek: G+C tartalmuk 55-67 mol%, így egy másik törzsbe, az Actinobacteria törzsbe tartoznak.
A Firmicutes törzs Bacilli osztályába és Lactobacillales rendjébe tartoznak a tejsavbaktériumok. Nem mozgó, nem spóraképző, kataláz-negatív, nitrátreduktáz-negatív, citokrómoxidáz-negatív, nem lélegző, aerotoleráns, igényes és savtűrő kokkuszok vagy pálcák. A szénhidrátok fermentációja során végtermékként tejsavat képeznek, nem folyósítják el a zselatint és nem termelnek indolt. A fenti általános jellemzés kivételeiként olyan fajok is előfordulnak a tejsavbaktériumok között, amelyek katalázt vagy citokrómokat képeznek hematin tartalmú táptalajokban (hem forrás lehet például a vér), illetve hemet nem tartalmazó, katalázt, pszeudokatalázt termelő fajok is vannak.
A tejsavbaktériumok egyedüli energiatermelő módja a tejsavas erjedés, működőképes teljes citromsavkörük, hemhez kötött elektrontranszport rendszerük és citokrómjuk sincs. Energiájukat a szénhidrátok szubsztrát szintű foszforilációjával nyerik. Következésképpen egyrészt savtűrők (szaporodásuk optimális pH-ja 5,5 körül van, de elviselik a jóval kisebb, 3,0-3,5-es pH-t is), másrészt tápanyagigényük összetett, saját szintézis hiányában különböző aminosavakra, fehérjékre, zsírsav észterekre, sókra, nukleinsav származékokra és vitaminokra van szükségük a szaporodáshoz.
Komplex tápanyagigényük miatt elsősorban olyan élőhelyeken fordulnak elő, ahol nagy mennyiségű oldott szénhidrát, fehérjebomlási termékek és vitaminok vannak jelen, vagyis növényi (gyümölcs, zöldség, gabona) és állati (tej, hús) eredetű anyagokon, erjesztett vagy romlott élelmiszerekben, emberi és állati szervezetek tápcsatornájában stb.. Mivel a tejsavbaktériumok sok fajának hosszú történeti kapcsolata van az élelmiszerekkel, ezért általánosan biztonságosnak (generally regarded as safe: GRAS) fogadjuk el őket.
A tejsavbaktériumok a tej legáltalánosabb mikroorganizmusai. Hasznosak, amikor színtenyészetek (pl. vajkultúra, joghurtkultúra, sajtkultúra) alkotóiként tejterméket állítunk elő velük, károsak, ha elszaporodva megsavanyítják a nyers vagy pasztőrözött tejet; sőt a csoport tartalmaz patogén baktériumokat is, amelyek nem kívánatosak az élelmiszerben (pl. több sztreptokokkusz, valamint a halpatogén karnobaktérium).
Az általuk végzett tejsavas erjedés két, lényegileg eltérő biokémiai úton is folyhat A glükóz homofermentatív erjesztése a glikolízis szerint történik, a piroszőlősav közvetlenül tejsavvá redukálódik. A glikolízist folytató sejtekben működik a fruktóz-difoszfát aldoláz.
A heterofermentatív tejsavas erjedés első szakasza más, a pentóz-foszfát utat követi. A sejtekből hiányzik a glikolízis kulcsenzime, az aldoláz és a triózfoszfát-izomeráz, de működik a foszfoketoláz, ami a glükonsavból képződő pentózokat hasítja. A heterofermentatív erjedés ezért egyrészt mindig gázképződéssel jár, másrészt végtermékei vegyesek és a fajok szerint változóak, köztük tejsav, ecetsav, etanol különböző arányban keletkezhetnek (valamint kisebb mennyiségben hangyasav és glicerin). Adott mennyiségű cukorból általában fele annyi energiát tudnak előállítani, mint a homofermentatívok.
Bár a tejsavas erjedésnek alapvetően kétféle mechanizmusa van, a tejsavbaktériumok erjesztési típusa háromféle lehet: obligát homofermentálók (pl. Lactococcus lactis), obligát heterofermentálók (pl. Leuconostoc mesenteroides), valamint fakultatív heterofermentálók (pl. Lactobacillus plantarum). Utóbbiak a glükózból csak tejsavat képeznek, de erjesztik a glükonsavakat és a pentózokat is (Deák, 2006).
A tejsavbaktériumok története és jelenlegi rendszertana
Joseph Lister 1873-ban számolt be először a tej savanyodásáért felelős mikroorganizmusok izolálásáról. A törzset Bacterium lactis-nak nevezte el, ezt később Streptococcus lactis-ra változtatták (Ward et al., 2003). Komoly hatással volt a tejsavbaktériumok rendszerezésére Sigurd Orla-Jensen monográfiájának megjelenése 1919-ben. Az általa használt főbb klasszifikációs tulajdonságok: morfológia (kokkusz vagy pálca), glükóz-fermentáció módja (homo- vagy heterofementatív), szaporodás néhány “kardinális” hőmérsékleti értéken (pl. 10°C-on és 45°C-on) és a hasznosított cukrok fajtája. Ahogy e fejezet végén látható lesz, ezek a tulajdonságok még mindig nagyon fontosak a tejsavbaktériumok osztályozásában. Orla-Jensen munkájának köszönhetően az a nézet terjedt el, hogy a tejsavbaktériumok csoportjának magját a Lactobacillus, Leuconostoc, Pediococcus és Streptococcus nemzetségek alkotják (Axelsson, 1998). Ezek a nemzetségek Lancefield 1933-ban javasolt szerológiai azonosítási rendszerében az N csoportba kerültek. Ez a szerológiai azonosítás elválasztotta őket az A, B, és C csoportba tartozó sztreptokokkuszoktól és a D csoportba tartozó enterokokkuszoktól (Ward et al., 2003). Schleifer és mtsai (1985) a korábbi Streptococcus nemzetséget először három részre osztották: Enterococcus, Lactococcus és Streptococcus sensu stricto. Azóta a tejsavbaktériumok osztályozása többször is jelentősen megváltozott (Axelsson, 1998).
A jelenlegi molekuláris filogenetikai osztályozás nem mindenben egyezik a hagyományos rendszertani csoportosítással, és új nemzetségek létrehozásával járt, amelyekhez a továbbiakban új fajokat is leírtak. A tejsavbaktériumokhoz tartozó nemzetségek 16S rRNS szekvencián alapuló konszenzus fáját az 1. ábra mutatja. A törzsfa szerint közeli rokon a Carnobacterium, az Enterococcus, a Vagococcus, az Aerococcus, a Tetragenococcus, a Lactosphaera és a Melissococcus nemzetség. Ugyancsak közeli rokonságban van a Lactococcus és a Streptococcus nemzetség, míg a Lactobacillus nemzetség filogenetikailag különálló ágat alkot. A Lactobacillus nemzetség genetikai heterogenitására utal, hogy a különböző fajok G+C tartalma nagyon széles, 32-53 mol% közötti tartományban helyezkedik el, míg általában, ha két faj között több mint 10 mol% a különbség, akkor már nem tartoznak ugyanabba a nemzetségbe (url1).
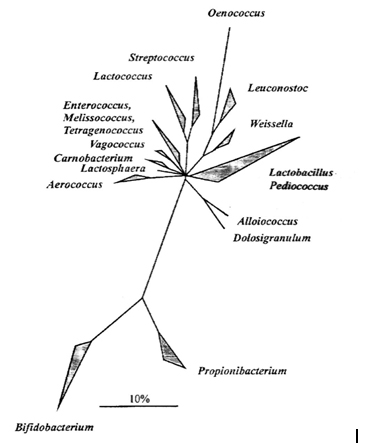
1. ábra: 16S rRNS szekvenciák összehasonlító elemzésén alapuló konszenzus fa; a vonal a filogenetikai távolságot jelöli (Holzapfel et al., 2001; url1)
A molekuláris szempontok szerint alkotott nemzetségeket csak részben jellemzik olyan közös alaktani és élettani bélyegek, amelyeket a tejsavbaktériumok korábbi osztályozásánál figyelembe vettek. A baktérium-taxonómiában a morfológiai jegyek kulcsfontosságú szempontként történő figyelembe vétele megkérdőjelezhető (Woese, 1987), ennek ellenére még mindig szerepet játszik a tejsavbaktériumok általánosan elfogadott jellemzésében (1. táblázat).

1. táblázat: A tejsavbaktérium nemzetségekre jellemző alaki és élettani tulajdonságok (Collins et al., 1993; Deák, 2006)
Tejsavbaktériumok antimikrobás anyagai
A tejsavbaktériumok antimikrobás hatását legtöbbször a szerves savak (Schillinger és Lücke, 1989) valamint a hidrogén-peroxid (Tagg et al., 1976; Gilliland és Speck, 1977) termelésének tulajdonítják. Mások szerint a szaporodásgátlás a termelt bakteriocinnek köszönhető (Klaenhammer, 1988).
A csökkent pH és a szerves savak
A fermentáció során a tejsavbaktériumok az alapanyagban található szénhidrátokat (elsősorban a glükózt és a laktózt) anaerob módon tejsavvá bontják, ezáltal a termék pH-ját a savas tartományig csökkentik (pH < 4,5). Ezt a savas közeget a tejsavbaktériumok többnyire jól tolerálják, néhány, romlást okozó és kórokozó baktérium viszont kevésbé viseli el. A savas kémhatás denaturáló hatással van a sejtfelszíni enzimekre, és a protonok citoplazmába való beáramlása miatt a sejt belső pH-ja is lecsökken, károsodásokat okozva a fehérjék és a DNS szerkezetében, a baktériumok anyagcsere folyamataiban.
A savas kémhatás mellett jelentős károsító hatásuk van a képződő gyenge savak (tejsav, ecetsav stb.) disszociálatlan molekuláinak is. Ezek a lipofil molekulák ugyanis könnyen átjutnak a plazmamembránon, és a citoplazmában disszociálnak. A sejtbe beszivárgó, valamint a disszociáció során felszabaduló protonok feldúsulnak a citoplazmában, és tönkreteszik a transzmembrán protongrádienst, ami szükséges a különböző transzportfolyamatokhoz, a mozgásképességhez és az ATP bioszintéziséhez. A baktériumok protonpumpák és ioncserélő csatornák segítségével, illetve negatív töltésű ionok felvételével igyekeznek helyreállítani homeosztázisukat, azonban ezek ATP-t igénylő folyamatok, amelyek előbb-utóbb kimerítik a sejtek energiatartalékait (Booth és Kroll, 1989). Egyes szerves savak (pl. hangyasav, ecetsav) disszociációjakor nem csak a felszabaduló protonok okoznak gondot, hanem a képződő anionok is, amelyek gátolják a baktériumok anyagcseréjét (Corlett és Brown, 1980; Szekér, 2007).
Hidrogén-peroxid
Oxigén jelenlétében a tejsavbaktériumok elektronokat visznek rá a molekulára, és ezáltal szuperoxid anion (O2-), hidrogén-peroxid (H2O2) vagy víz (H2O) keletkezik. A hidrogén-peroxid erős oxidálószer lévén képes gátolni, illetve elpusztítani a romlást okozó és a patogén baktériumokat (Szekér, 2007).
Bakteriocinek
A tejsavbaktériumok szerves savak termelése mellett fehérje természetű antimikrobás anyagok, ún. bakteriocinek termelésével is képesek gátolni más, elsősorban Gram-pozitív mikroorganizmusok szaporodását. Tagg és mtsai (1976) definíciója szerint a bakteroicinek fehérje jellegű vegyületek, amelyek közeli rokonságban levő baktériumokat képesek elpusztítani. Bár ez a meghatározás a bakteriocinek többségére igaz, ismertek olyanok is, amelyek rendszertanilag távolabbi baktériumcsoportok ellen is hatásosak, és a fehérjerész mellett lipid- illetve szénhidrát-komponenseket is tartalmaznak (Marugg, 1991; Barefoot és Nettles, 1993).
A számtalan felfedezett tejsavbaktérium bakteriocin közül a Lactococcus lactis subsp. lactis által termelt nizin (E234) az egyedüli, amelynek élelmiszer-tartósítószerként való felhasználását a WHO engedélyezte. A nizin viszonylag széles hatásspektrummal rendelkező antimikrobás anyag, amely a Gram-pozitív baktériumok sokaságát gátolja (Delves-Broughton, 1990), úgymint néhány Staphylococcus, Enterococcus, Pediococcus, Lactobacillus, Leuconostoc, Listeria, Corynebacterium fajt, valamint a Mycobacterium tuberculosis-t. A legnagyobb jelentősége abban rejlik, hogy a Bacillus és a Clostridium spórák kicsírázását is gátolja. Ez utóbbi tulajdonságát a tejipar hasznosítja oly’ módon, hogy a nizint a sajtgyártás alapanyagához, a tejhez adagolja, meggátolva ezzel a sajtok Clostridiumok okozta késői puffadását.
Magyarországon elsőként Pulay (1954) számolt be a nizin tejipari alkalmazásának lehetőségéről, a sajtgyártás során előforduló vajsavas puffadást gátló hatását hangsúlyozva. Munkatársai segítségével számos üzemi kísérletet végzett nizinnel, illetve nizin-termelő törzsekkel sajtok vajsavas puffadásának megakadályozására (Pulay et al., 1956). Különösen jó eredményeket kaptak röglyukas félkemény sajtok (pl. Óvári sajt) esetében.
A nizin vegetatív sejtekkel szembeni antimikrobás hatása abban rejlik, hogy a citoplazma-membránba beépülve azon pórusokat hoz létre, amelyeken keresztül kiegyenlítődik a membránpotenciál kialakításában szerepet játszó ionok koncentrációja a membrán két oldalán, megszüntetve ezzel a protongrádienst. A nizinmolekulák összekapcsolódva alakítják ki a membránt átívelő csatornákat. A nizin hatását segíti a közeg savas kémhatása, hiszen a jelentősebb proton koncentráció-különbség meggyorsítja a hidrogénionok kiegyenlítődést a membrán két oldalán. A kisebb pH emellett azért is fontos, mert savas környezetben megnő a nizin oldhatósága és stabilitása, lúgos közegben pedig inaktiválódik a molekula (Garcerá et al. 1993). A nizin nem jelent veszélyt az emberi szervezetre, mert a bélcsatorna emésztőenzimjei (α-kimotripszin) gyorsan inaktiváják (Szekér, 2007).
A tejsavbaktériumok szerepe a fermentált élelmiszerek előállításában
A tejsavasan erjesztett élelmiszerek alapanyaga lehet tej, hús, zöldség vagy gabona, amelyekből a fermentáció körülményeinek szabályozásával változatos élelmiszerek készíthetők. Az így készült termék – a tejsavbaktériumok tevékenységének eredményeképpen – az alapanyaghoz képest változatosabb, biztonságosabb, jó minőségű, hosszabban eltartható, íz- és tápanyagokban gazdagabb lesz, ugyanakkor az esetlegesen jelenlevő antinutritív anyagok mennyisége csökken. Ilyen élelmiszerek a fermentált tejtermékek (pl. joghurt, kefir, tejföl, vaj, sajtok), a fermentált húskészítmények (pl. szalámi- és kolbászfélék), az erjesztett zöldségfélék (pl. savanyú káposzta, uborka, olívabogyó, kávébab, kakaóbab) vagy a savanyú kovászos kenyér (Deák, 2006; Galántai, 2008).
Az erjesztés a kezdetekben spontán fermentációval történt, az alapanyagban lévő vagy a környezetből véletlenszerűen belekerülő baktériumok segítségével. Ezt a módszert alkalmazzák ma is a zöldségfélék tartósításánál. Később az előző erjesztésből megmaradt baktériumtömeg továbboltásával igyekeztek azonos minőségű terméket létrehozni. A mikroorganizmusok a folyamatos átoltással való fenntartás során alkalmazkodtak a különböző alapanyagokhoz, így ezekben jól és gyorsan el tudtak szaporodni. A baktériumok felfedezése és a mikrobiológiai módszerek fejlődése tette lehetővé a fermentációban részt vevő fajok megismerését, pontos jellemzését és ezeknek az ismereteknek a birtokában a starterkultúrák kifejlesztését.
A tejipar számára a tejsavbaktériumok közül a következő nemzetségek jelennek meg kultúraalkotóként: Lactococcus (Lc.), Enterococcus (Ec.) Lactobacillus (Lb.), Leuconostoc (Ln.), Pediococcus (Pc.) és Streptococcus (Sc.) (International Dairy Federation, 1996).
A Galen Bio Kft. akkreditált mikrobiológiai laboratóriuma vállalja a Mezofil tejsavtermelő baktériumok számának meghatározást hagyományos tenyésztéses módszerekkel.

Tejsavtermelő baktériumok kimutatása MRS táptalajon